As I was looking at my summer schedule, it occurred to me that my time out in the field (here defined as outside, collecting data, probably wearing zip-off pants and carrying a field book, insect net and a set of vials) has been getting less and less, every year. As a PhD student I spent most of my summer collecting data. I loved it - the rugged joys of bumpy back-roads in Alberta, the sticky and smelly combination of sweat and bug spray, the cold beer at the end of a long field day. As I moved on to a post-doc in Ohio, I still spent a lot of time collecting spiders in soybean fields, helping other graduate students in the field, although the summers also included some lab work, and substantial time writing manuscripts.
When starting at McGill over 10 years ago, I kick-started my research program by spending weeks in the field, and seemed to manage a lot of time with each of my graduate students during the field season. However, time in the field was measured as weeks, and not months. Now, as I look at my schedule, I’m “maybe” going to get one full week in the field this summer, and a fews days here and there helping with other projects going on in the lab. My time doing field work, actively collecting data, is minimal.

Deep thoughts: field work in the Arctic. Are these days long gone…?
Wait a second. One reason I got into this business was because I like to figure out neat stuff about nature, while being in nature. As a child, I always enjoyed being ‘in the field‘ (this is also known as ‘playing outside‘) and wanted to continue this as an adult. What happened?
Academics in my discipline of study (let’s call it ‘field ecology‘) and at my career stage (i.e., some years into the job) spend relatively little time in the field and the bulk of their time is a desk job, click-click-clicking away on a keyboard. Staring at a monitor. I know there are exceptions (and BIG congratulations on those of you who do manage to get outside to collect data, regularly!), but when I look around to my colleagues, most of them spend more looking out a window instead of being out that window. The time gets chewed up by other (important) priorities: grant writing, editing manuscripts, writing manuscripts, answering emails, reading papers, attending meetings, chairing meetings, going to conferences, preparing talks for those conferences, writing lectures, delivering lectures and so on. These are all the current demands on our time, and they are the things that the job requires! (for other relevant discussions about this, have a peek at this post by Sarah Boon, and I’ve previously written about how I spend my time).
Bottom line: most of my work duties are indoor activities. I am fortunate in that some of my teaching occurs outside, but that is not the norm. The other thing that happens is ‘life’ - time with family is important to me, and time away from family is difficult. One reason I’ve spent less weeks away is because it’s tough on all of us and I like being around when the kids are growing up. There’s also that thing called a vacation - Academics typically their vacation time during the summer. (related to this is a post over at Dynamic Ecology titled “how often do you travel”, by Meg Duffy)
That is how I have traded field biology for a desk job.
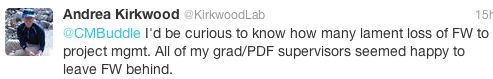
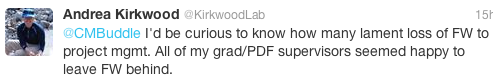
I’m not alone: here are some responses from folks on Twitter when I asked about their experiences, and whether they have traded field biology for a desk job:




This is not a lament; this is not a sob story. In fact, perhaps many of us are OK with this transition from field biologist to ‘research manager’:
There is an important message here for people moving up through the Academic system: current PhD and MSc students need to recognize that the idea of landing an Academic job that gets you ‘out in the field’ a lot is probably a pipe dream.
I’ll end with some optimism: Even though things have changed, I think I can still call myself a ‘field ecologist’ and here’s why:
1) “Field Trips” can be short. It’s possible to capture an hour outside over lunch and collect data on Agelenopsis spiders in a hedge near the picnic table, or stop off at a bird banding station in the AM before work, or swing by a forest to check a pitfall trap on the way home. I have come to realize that field work need not be ‘weeks away’. In many cases, it’s worth starting up a project that takes you outside regularly, at a local field site. This makes the field work an easier part of the day and you don’t need to schedule weeks away (nor will you need to schedule it months in advance). Keeping it simple, and keeping in manageable is important for me, given the other constraints on time.
2) Trade-offs: I spend time in the field instead of attending a lot of conferences. I have always enjoyed going to scientific conferences, but given the difficulties in getting away for extended periods of time, I realized that I could do field work, or attend conferences, but doing both is not always possible. One of my academic mentors discussed this with me soon after I had started my job at McGill (ironically, at a conference!); he said that when the weather is good, time was better spent collecting flies rather than sitting in a hotel basement. Good point. (By the way, summer-conference are kind of annoying because of this conflict!). Networking at conferences is very valuable, but that face-to-face networking may not be as essential later in a career. Thanks to social media, it’s also possible to attend conferences virtually.
3) Live vicariously through students: My thoughts about field work are somewhat nostalgic and dreamy, and I forget about the problems. I forget about the flat tires, encounters with bears, the biting flies, and the exhaustion. I’m reminded of these things when my graduate students come back from the field, and sometimes I am happy I wasn’t with them. I can, instead, spend a day or two with them in the field, troubleshoot, help but not have to suffer through it all. I’m a ‘gentleman field biologist’ now. Is that lame? Is that pathetic? Nope. I’ve put in my time and can now have my field trips field with all the fun parts and less of the annoying parts.
4) Mixing vacations with field biology: I’ve not been all that successful at this, but I do know colleagues who manage to mix extended vacation time with field work. I do this on a smaller scale, and it typically includes carrying vials along with every trip, whether it is to the family cottage, or just a walk in the local forest. I’m always after records of pseudoscorpions, and have managed some nice finds while on vacation. My family does, however, gives me strange looks when I go chasing after spiders or butterflies during lunch break while on a road trip. I can handle the ridicule -> it’s for science!
Although I have largely traded field work for a desk job, there are still glimmers of exciting field work, and still opportunities to get outside and be reminded of the reasons why I originally got into this line of work. I am not depressed or sad about my desk job - I have the best job in the world, despite the the fact that I stare out the window and sometimes dream of field work. I also maintain that these things come in cycles - a few years ago I was away for a few weeks in the field, even if this year is less intensive. It’ll come around again, and perhaps I will write a post in the future that discusses how it’s possible to be a gritty, smelly, rough and tough field biologist again. For now, though, I must stop typing. It’s hard work and my fingers are a little sore.