This post is written by PhD student Shaun Turney, and highlights a recent publication from the lab.
Two years ago, I was finishing my MSc and considering whether I’d like to do a PhD, and if so, with whom. I met with Chris and we threw around a few ideas for PhD projects. It was when he brought up a certain mystery that my decision to do a PhD in his lab was cemented. The mystery? Chris and his former PhD student Crystal Ernst were puzzled why there seem to be so many carnivores on the Arctic tundra, and relatively few herbivores to feed them.
How could it be possible? Is there a high level of cannibalism? (But then it would be like pulling oneself up by ones bootstraps — how does the energy and biomass enter the carnivore population in the first place?) Are the carnivores really omnivores? Is our methodology for sampling the tundra biota biased towards carnivores? Is the transfer of energy from herbivores to carnivores somehow more efficient (less energy loss) than in other ecosystems? These sorts of questions touch on some fundamental questions in ecology and I was hooked.
Shaun Turney, vacuuming the Tundra.
It seemed to me the logical first step would be to find out what is a typical predator-prey ratio. In what proportions are the organisms in an ecosystem divided up from plant (lowest trophic level) to top predator (highest trophic level)? The answer to that questions has already been very much explored when it comes to biomass and abundance. Charles Elton explained about 80 years ago that typically the mass and number of organisms form “pyramids”: They decrease with trophic level because energy is lost with each transfer from resource to consumer. But what about diversity? How does the number of species change with trophic level?
I decided to look at the food webs in the data base GlobalWeb to answer this question, and we just published a paper in Oikos on this topic. I found that typically ecosystems form “pyramids of species richness”, just like the pyramid of numbers and pyramid of biomass described by Elton. But some types of ecosystems, notably in terrestrial ecosystems, we can consistently observe a uniform distribution or even an “upside-down pyramid” rather than a pyramid like Elton described. That is, there are consistently cases where there more carnivore species than herbivore species in an ecosystem.
An example of aquatic compared to terrestrial food-web structure (from Turney and Buddle)
So evidently, at least when it comes to diversity, the pattern that Chris has observed in the tundra is not so unusual! The next step for me is to try to figure out why. Stay tuned!
Reference:
Turney S and CM Buddle. Pyramids of species richness: the determinants and distribution of species diversity across trophic levels. Oikos. DOI: 10.1111/oik.03404
This is the second post by Honour’s undergraduate student Kamil Chatila-Amos - he has been busy working on identifying LOTS of spiders from bogs of northern Quebec. His first blog post introduced his project: this one gives a glimpse into the data…
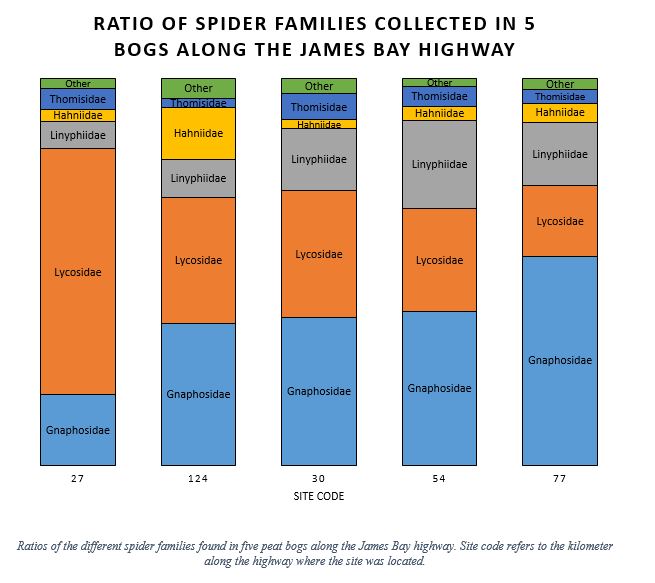
My project is focused on studying spiders from bogs in the James Bay region of Quebec. Five bogs along the James Bay highway were sampled with pan traps every week for four sampling periods. In the full project I’m looking at how abiotic factors (i.e. pH, water table, latitude, etc.) and the plant community affect the arachnid community composition. For now, let’s look at how the spider families are distributed in these sites:
The first thing that might strike you if you are familiar with the area and its spider fauna is that in 4 out of 5 sites, neither Lycosidae (wolf spiders) nor Linyphiidae (subfamily Erigoninae) are the most abundant family. Previous studies in similar habitats tend to find a much greater proportion of those two taxa (Aitchison-Benell 1994; Koponen 1994). All sites except the first have more Gnaphosids than Lycosids. However, the breakdown within families is very different. Whereas the Lycosids are represented by 19 species, there were only five species within the Gnaphosidae. Even more impressive is that one Gnaphosidae species represents 99% of the family. Indeed, Gnaphosa microps alone represents a fifth of all arachnids I collected.
I’ve come to like Gnaphosa microps a lot! The family Gnaphosidae is pretty easy to identify thanks to their long and separate spinnerets, colour and eye placement. Even the palps, which are unique to species, are fairly easy to recognize. It ranges in size from 5.4 – 7.1 millimeters which is a large enough size so it isn’t a hassle to manipulate.
Gnaphosa microps is by no means a star of the spider world but we still know a fair bit about it. It is a holarctic species meaning it can be found in almost all of the northern hemisphere, even as far as Turkey (Seyyar et al. 2008). It is usually found in in open boreal forests, alluvial meadows and bogs. A nocturnal species, it spends its days in a silk retreat under moss or debris and hunts at night by catching prey on the ground (Ovcharenko et al. 1992). Even though sampling has been done very near my sites and in similar habitats (Koponen 1994) I still haven’t found another study where it was the most abundant species.
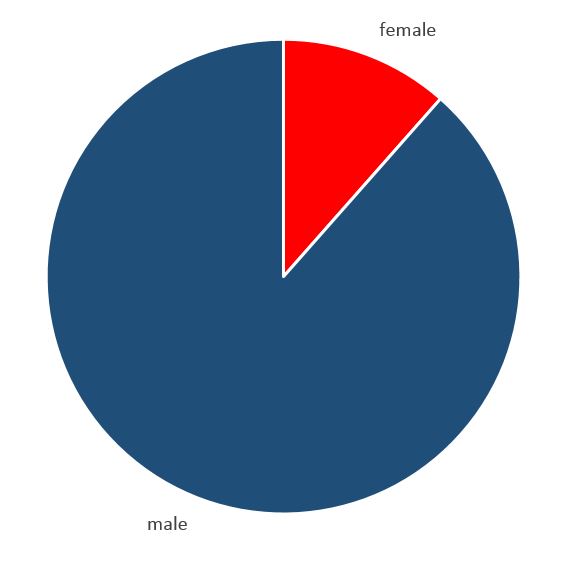
Another interesting tidbit about this species is just how skewed their sex ratio is. According to my data, males outnumber females almost 10 to 1! Now this does not mean it is always like this in nature, this ratio can be explained by sexually dimorphic behavior. This means that the males would behave differently than females in a way that would increase their odds of falling into traps. Indeed, according to Vollrath and Parker (1992) spider species with sedentary females have smaller, roving males. And like their model predicts the G. microps males are a bit smaller than the females.
Sex ratio of Gnaphosa microps, collected in bogs
So what’s next? I still need to retrieve the COI barcode of all my species and that will be possible thanks to the University of Guelph’s Biodiversity Institute of Ontario. This is to make sure my identifications are indeed correct. As a first time spider taxonomist it’s great to be able to confirm my work in a way that still is not widely available. Today I received the plate in which I’ll load the spider tissue and I am amazed at how tiny it is. I guess they just need 2mm per spider but I still expected it to be much more impressive. Hopefully I don’t get any nasty surprises once the DNA data comes back, although some of those tiny Linyphiids did give me a pretty bad headache…
Ovcharenko VI, Platnick NI, Sung T. 1992. A review of the North Asian ground spiders of the genus Gnaphosa (Araneae, Gnaphosidae). Bull. Am. Museum Nat. Hist. 212:1-92
I’m excited to announce a recent paper to come out of the lab, by former PhD student Dorothy Maguire, and with Dr. Elena Bennett. In this work, we studied the amount of insect herbivory in forest patches in southern Quebec: the patches themselves varied by degree of fragmentation (ie, small versus large patches) and by connectivity (ie, isolated patches, or connected to other forest patches). We studied herbivory on sugar maple trees, both in the understory and canopy, and at the edges of the patches. Our research is framed in the context of “ecosystem services” given that leaf damage by insects is a key ecological process in deciduous forests, and can affect the broader services that forest patches provide, from supporting biodiversity through to aesthetic value. Dorothy’s research was part of a larger project about ecosystem services and management in the Montérégie region of Quebec.
Dorothy Maguire sampling insects in the tree canopy (Photo by Alex Tran)
The work was tremendously demanding, as Dorothy had to select sites, and within each site sample herbivory at multiple locations, including the forest canopy (done with the “single rope technique). Dorothy returned to sites many times over the entire summer to be able to assess trends over time. Herbivory itself was estimated as damage to leaves, so after the field season was completed, thousands of leaves were assessed for damage. The entire process was repeated over two years. Yup: doing a PhD requires a suite of skills in the field and lab, and there is no shortage of mind-numbing work… Dedication is key!
As with most research, we had high hopes that the results would be clear, convincing, and support our initial predictions - we certainly expected that forest fragmentation and isolation in our study landscape would have a strong effect on herbivory - after all, our study forests varied dramatically in size and isolation, and herbivory is a common and important ecological process, and insect herbivores are known (from the literature) to be affected by fragmentation.
The landscape of southern Quebec. Lots of agriculture, some patches of forest.
However, as with so much of ecological research, the results were not straightforward! “It’s complicated” become part of the message: patterns in herbivory were not consistent across years, and there were interactions between some of the landscape features and location within each patch. For example, canopies showed lower levels of herbivory compared to the understory, but only in isolated patches, and only in one of the study years! We also found that edges had less herbivory in connected patches, but only in the first year of the study. Herbivory also increased as the season progressed, which certainly makes biological sense.
So yes, it’s complicated. At first glance, the results may appear somewhat underwhelming, and the lack of a strong signal could be viewed as disappointing. However, we see it differently: we see it as more evidence that “context matters” a great deal in ecology. It’s important not to generalize about insect herbivory based on sampling a single season, or in only one part of a forest fragment. The story of insect herbivory in forest fragments can only be told if researchers look up to the canopy and out to the edges; the story is incomplete when viewed over a narrow time window. In the broader context of forest management and ecosystem services, we certainly have evidence to support the notion that herbivory is affected by the configuration of the landscape. But, when thinking about spatial scale and ecosystem processes, careful attention to patterns these processes “within” forest patches is certainly required.
We hope this work will inspire others to think a little differently about insect herbivory in forest fragments. Dorothy’s hard work certainly paid off, and although the story is complicated, it’s also immensely informative and interesting, and sheds light on how big landscapes relate to small insects eating sugar maple leaves.
Reference:
Maguire et al. 2016: Within and among patch variability in patterns of insect herbivory across a fragmented forest landscape. PlosOne DOI: 10.1371/journal.pone.0150843
This is the second post by undergraduate student Jessica Turgeon - she’s doing an Honour’s project in the lab; here’s her first post that introduces the project. Since that first post, Jessica has spent a LOT of time at the microscope, and has identified over 120 species of spiders and beetles from forest canopies and understory habitats.
Every species has a different story to tell and each one of these is equally interesting. I sometimes think about natural history as ‘fun facts’: something interesting about an organism (or species) to tell children so that they can appreciate nature. As my time at McGill progressed and my knowledge of the natural world deepened, I realized that the ‘fun facts’ are actually built upon a very strong scientific foundation, and can help us understand results of research projects. Natural history can sometimes be reduced to ‘fun facts’ but it’s a whole lot more than that!
The European Snout Beetle on a pin.
I decided that perhaps I should look at the natural history of some of my species and maybe this would shed light on some patterns that I’m seeing within the data. The most abundant beetle species was Phyllobius oblongus (Curculionidae) with 69 individuals. Interestingly, we only collected this species in the first half of our sampling season and they were mainly collected on black maple and sugar maple trees. To try and understand why this is so, I turned to the species’ natural history, and to the literature.
These weevils tend to eat fresh leaf shoots and prefer the soft leaves found on maple trees. Once the maple’s leaves are fully-grown, P. oblongus moves on to plants with indeterminate growth, like raspberry bushes (Coyle et al. 2010). This corresponds exactly to our data: the beetles were found on our black and sugar maples during the beginning of summer and then they taper off as the season progressed!
Beetle data: the European Snout Beetle was only collected during the beginning of the season.
To make this even more interesting, P. oblongus is an invasive species. Its common name is the European Snout Beetleand was accidentally introduced into North Eastern North America in the early 1900s. While most invasive species are a cause for concern, both the Canadian and American governments largely ignore this species. It may inflict some damage to trees but not enough to be worried about. They’re more annoying to researchers than anything since they congregate in the trees in large numbers!
The second most abundant beetle species in the collections was Glischrochilus sanguinolentus (Nitulidae). This species is native to Canada and rather abundant. Species in this genus are called sap beetles but this species in particular is more commonly called a picnic beetle. Large groups of G. sanguinolentus swarm to picnics since they are attracted to sweet food, which ruins the picnics. In nature, they feed on the sap produced by injured trees - hopefully not an indication that the trees we were climbing were damaged!
The natural histories of species open new doors to understanding how organisms live and interact with one another. I thought that it was strange that P. oblongus completely disappeared from my samples midway through the sampling season and its natural history explained why this was so. Picnic beetles eat the exuding sap of an injured tree so in the future I’ll be on the lookout so that I don’t accidentally climb a broken tree! So really, natural history is more than just ‘fun facts’; it helps us understand patterns and to better understand how our natural world works.
This is a guest post, written by an Honour’s undergrad student in the lab, Kamil Chatila-Amos. It’s the first of two posts about his work, and the goal of this post is to introduce Kamil and his research project.
Research can be serendipitous and spontaneous, and that’s certainly the story of how my honour’s project started! I spent last winter working on howler monkeys in Panama (which is a story in itself) and although I adored every second of it, it certainly made me out of touch with the McGill world. When I came back, most of my friends had found themselves a summer research job and even an honours supervisor for the upcoming semester.
So there I was, barely a week after my return, erratically filling out online job applications in the lobby of one of our buildings. I was looking at all kinds of opportunities: herbarium employee in Edmonton, ichthyology assistant in Wisconsin, plant surveying in Vaudreuil, bird surveys in Ontario, insectarium employee in Montreal. I was applying to anything and everything that was still available. Little did I know that the arthropod ecology lab is right next to the lobby… Chris walked by, saw me and stopped to chat. (Well it’s more accurate to say he talked to me while quickly walking to his next meeting)*. Turns out, a student of Terry Wheeler (another entomology Prof. at Macdonald campus), Amélie Grégoire Taillefer, was going to post a job online that very afternoon! She was looking for a field assistant to help her catch flies in bogs in the James Bay area.
A couple days later I was northern-bound! A 15 hour drive north of Montreal is the town of Matagami and about 30 km north of there is Lake Matagami, along which we were staying. In a yurt. A yurt!!! Basically, a large round tent of Mongolian origins. They’re big and this one had a minimal kitchen and shower. But the fact remains that it’s a tent with the isolative properties of canvas. It got pretty cold those first couple weeks and dropped below freezing a few nights. At least it had a fireplace. (It’s actually a great place for people wanting to explore that area of Québec and the owners are wonderful. Go check them out at ecogiteslacmatagami.ca)
The work itself was great. The first week, we explored the area for suitable bogs to install her pantraps. That’s when I realized how awesome bogs are. There are so many things to eat in bogs! Cattails, cranberries, Labrador tea, cloud berries, chanterelles, boletes, black flies…
For the remainder of the trip two days a week were spent visiting our five sites and harvesting the pantraps filled with flies, dragonflies, crickets, spiders and the occasional putrid mouse. The following two or three days we would sort through the samples, separating the lower flies (Nematocera) from the rest.
Ready for some serious bog-sweeping.
After the first week I couldn’t help but notice just how many spiders we were catching. Mostly out of pity I think, I decided to sort out the spiders as well. I felt bad throwing them out… Fast forward to five weeks later and I’m heading back to Montreal with a bagful of vials filled with dead spiders. (My roommates were not very fond of having them in our freezer).
A few weeks later I set up a meeting with Chris and essentially barged into his office with the spiders to ask to work in his lab. It took a while (and quite a bit of convincing) but here I am, sorting through spiders and writing blog posts!
The research project we structured has two components. The first part will look at how the community composition of spiders varies between the five sampled bogs. Second, I’m lucky enough to have the opportunity to try DNA barcoding using COI markers. This part remains very blurry right now**, but I’m very excited to see where it leads.
Kamil hard at work in the lab!
If it weren’t for serendipity I would not have gone to James Bay this summer. And if it weren’t for being spontaneous, I would not have sorted out the spiders and would not be working in Chris’ lab right now. But spontaneity does have its down sides. I didn’t plan far enough ahead** and in hindsight, I should have collected some insect orders to be able to do a more in depth ecological analysis.
—-
* um, yes, I spend a LOT of time in meetings, and often have discussions and chats with student on my way to and from those meetings!
** for what it’s worth, research is often blurry, and planning ahead isn’t always possible!
This year I have the pleasure of teaching my Population and Community Ecology class in one of McGill’s Active Learning Classrooms - this one is touted as been quite exceptional, and I’m keen to put it to the test. Over the past 4-5 years, I have been teaching my quantitative ecology course almost entirely with chalk. In fact, I have actively argued about the value of teaching with chalk, and about a move away from technology can be beneficial to student learning, to my own teaching, and overall a very positive experience for all. Now I’ll be faced with this kind of environment when teaching my class:
A view of two of the group tables in the Macdonald Campus active learning classroom; each “pod” seats 12 students (in three wings), and each is colour coded, and linked and adjacent to a screen and whiteboard
So why change?
One problem with the Chalkboard it that it doesn’t easily allow for ‘capturing’ the content from the board. Students need to write their own notes (which is good, in my opinion), but at times there may be specific equations, graphs, or other content from the board that they wished they could have captured, but failed to do so. The Active Learning classroom allows an easy way to overcome this - as the tablet/screen that is in the room has a smart pen, and acts like a chalkboard (or, rather, kind of like a smartboard, except that the instructor uses the screen at the podium in the middle of the room). I can therefore project this board, and teach as if I was using chalk, and everything I write is projected on one of the screens. The big benefit here is that I can save everything I write as a PDF (or other file type), and upload the notes to the online course management system. This approach still encourages students to come to class and take notes, but doesn’t put them in a position to rush with notetaking, and live in fear of missing something that I write on the board. Here’s an example from the first lecture (it’s a bit clunky, and I’m not used to writing on the screen yet, but hopefully will get more seamless as the term progresses):
Another great benefit of the classroom is that it allows a second screen to be projected simultaneously as the first screen - I am thinking of primarily using this second screen to project graphs or equations directly from the textbook, through the use of a very high quality document camera. This saves me from having to hand-draw graphs (I do this very, very poorly - drawing straight lines is NOT easy!), and will allow the students to see the very direct ways that the content relates to the course’s textbook. This photo below shows how this looks: in that example the textbook cover is projected on the left screen and some ice-breaking questions are presented on the right-hand screen.
I have been trying to transition my course into more of an active learning course, and set an active learning challenge last year. Overall I felt this was very beneficial, but the traditional lecture theatre (where I have taught in the past) is not conducive to active group work and student collaboration. I’m excited that the active learning classroom is ideally set up for this: the 84 students in the class sit at seven separate tables, each with 12 students, and the tables are designed into three wings of four students each. This is optimal for group work, and provides many opportunities for different sizes of groups. Next to each table is a whiteboard and screen, and each table can project (independently) onto their screen. Students then have many options to collaborate and work on problems. I’m excited about this, and look forward to having groups of students work on problems together, collaboratively. The interesting thing about this space is that it doesn’t seem that big, yet is a classroom that holds 84 students! It’s also designed so that the noise level isn’t overwhelming when students are working in groups.
Students writing out ideas/answers to some questions, with their groups (from two different groups - notice the different colours?)
During the first lecture last week, I asked students how many of them had previously taken classes in the Active Learning classroom, and of those that did, many stated they did not enjoy the classroom. A little more discussion revealed that the students who disliked the classroom said their instructor used the room as a traditional lecture hall, and taught with powerpoint slides, from a podium. This clearly doesn’t work - the podium is in the middle, there are multiple screens (students say they are confused about where to look), and there are pillars that run right through the classroom (unavoidable since these are support pillars - and the classroom is in a basement); the ‘feel’ of the room, when used for traditional podium lectures, is all wrong. To use an Active Learning classroom means moving away from a podium-style lecture.
I certainly have a challenge ahead: in order to fully use the classroom’s potential, and make it engaging for the students, I have to ensure the technology and space is used properly. I’m only at the very start of term, so I will certainly report back on the experiences as the course unfolds. That being said, the combination of the technology and design is really promising for an active learning environment for my ecology class. It may just lead to the abandonment of chalk for a smart pen…
Note: this post is written by undergraduate Honour’s student Jessica Turgeon, who is a member of the arthropod ecology laboratory. This post is part of the requirements for her project, and is an introduction to her research.
I’ve always been interested in nature and the environment but was never a big fan of insects. As time went on and I learned to appreciate all organisms big and small I realized that I didn’t really have a preferred “pet taxon” but rather was interested in ecology and community structure. I found others that my interests were shared with other members of the arthropod ecology lab, and I was able to start an Honour’s project in the lab earlier this fall.
Using a beat-sheet in the tree canopy, to collect arthropods
I was given an opportunity to do an internship at Kenauk Nature, a 65,000-acre plot of land near Montebello, Quebec. This property is primarily used for the hunting and fishing industries, but they are branching into scientific research. Kenauk was keen to support three McGill interns to complete the Black Maple project, the pilot project for Kenauk Institute.
The Black Maple project revolves around black maples, since Kenauk is the only area in Quebec to have a black maple stand. The project consisted of three sub-projects, one for each intern and each project dealing with a different taxon. While the two other students worked on plants and birds, my project was about arthropods and their diversity in Kenauk. We wanted to characterise the community structures of beetles and spiders based on vertical stratification and tree species: this involved tree-climbing!
Jessica - getting ready to climb up!
During the summer, I looked at abundance data and concluded that beetles were more abundant in the upper canopy and that spiders were more abundant in the understorey. This internship transitioned into my Honour’s project, where I plan to look at species richness and functional diversity to answer my questions on community assemblages. To my knowledge, this has never been done at Kenauk Nature and would provide great baseline data for the owners of the property.
We sampled in three sites, each containing three trees. Each site had one sugar maple (Acer saccharum), one black maple (Acer nigrum) and one American basswood (Tilia americana). Within each tree we sampled five times: twice in the understorey, once in the middle canopy and twice in the upper canopy. We also used two different types of traps: beat sheets, an active technique, and Lindgren funnels, a passive technique. Both trap types are specialized, with beating more tailored towards spiders and Lindgren funnels invented to collect beetles. When beating a branch, the arthropods fall on a 1m2 sheet and are then collected whereas Lindgren funnels are hung in a tree and passively collect arthropods that fly into it.
As part of our job, we learned how to use a single ropes climbing system, a one-person method of using ropes to climb a tree. All three interns caught on quickly and it easily became our favourite part of the job. However, we did have to sort through the samples, a job requirement that wasn’t nearly as fun as climbing trees. But this is what happens in ecology: you romp around in the woods to collect your data then spend time in the lab analysing them. It was nice to experience this first-hand and I must say, I liked it and am looking forward to future projects like this.
Now that the summer is over and collection is completed, I spend all of my free time in the lab identifying beetles and spiders. All of the beetles are identified and about half of the spiders are identified. From this work, Kenauk Nature can proudly say that the property supports 24 families representing 117 species of beetles! Once the Kenauk Institute officially launches, more rigorous research can be done to try and increase these numbers.
Learning Taxonomy… spider drawings (of male palps) help.
All in all, from the sampling in the summer to the identification in the lab, this has been a great experience. Here’s to hoping the second half of my honours project will be as equally fun and challenging as the first half was! Stay tuned for a blog post to be published in the spring of 2016: it will summarize the main results from this Honour’s project.
Ecological monitoring is an important endeavour as we seek to understand the effects of environmental change on biodiversity. We need to benchmark the status of our fauna, and check-in on that fauna on a regular basis: in this way we can, for example, better understand how climate change might alter our earth systems. That’s kind of important.
A northern ground beetle, Elaphrus lapponicus. Photo by C. Ernst.
With that backdrop, my lab was involved with a Northern Biodiversity Program a few years ago (a couple of related papers can be found hereand here), with a goal of understanding the ecological structure of Arthropods of northern Canada. The project was meant to benchmark where we are now, and one outcome of the work is that we are able to think about a solid framework for ecological monitoring into the future.
A few weeks ago our group published a paper* on how to best monitor ground-dwelling beetles and spiders in northern Canada. The project resulted in over 30,000 beetles and spiders being collected, representing close to 800 species (that’s a LOT of diversity!). My former PhD student Crystal Ernst and MSc student Sarah Loboda looked at the relationship between the different traps we used for collecting these two taxa, to help provide guidelines for future ecological monitoring. For the project, we used both a traditional pitfall trap(essentially a white yogurt container stuck in the ground, with a roof/cover perched above it) and a yellow pan trap (a shallow yellow bowl, also sunk into the ground, but without a cover). Traps were placed in grids, in two different habitats (wet and “more wet”), across 12 sites spanning northern Canada, and in three major biomes (northern boreal, sub-Arctic, and Arctic).
Here’s a videoshowing pan traps being used in the tundra:
Both of the trap types we used are known to be great at collecting a range of taxa (including beetles and spiders), and since the project was meant to capture a wide array of critters, we used them both. Crystal, Sarah and I were curious whether, in retrospect, both traps were really necessary for beetles and spiders. Practically speaking, it was a lot of work to use multiple traps (and to process the samples afterwards), and we wanted to make recommendations for other researchers looking to monitor beetles and spiders in the north.
The story ends up being a bit complicated… In the high Arctic, if the goal is to best capture the diversity of beetles and spiders, sampling in multiple habitats is more important than using the two trap types. However, the results are different in the northern boreal sites: here, it’s important to have multiple trap types (i.e., the differences among traps were more noticeable) and the differences by habitat were less pronounced. Neither factor (trap type or habitat) was more important than the other when sampling in the subarctic. So, in hindsight, we can be very glad to have used both trap types! It was worth the effort, as characterizing the diversity of beetles and spiders depended on both sampling multiple habitats, and sampling with two trap types. There were enough differences to justify using two trap types, especially when sampling different habitats in different biomes. The interactions between trap types, habitats, and biomes was an unexpected yet important result.
Our results, however, are a little frustrating when thinking about recommendations for future monitoring. Using more than one trap type increases efforts, costs, and time, and these are always limited resources. We therefore recommend that future monitoring in the north, for beetles and spiders, could possibly be done with a trap that’s a mix between the two that we used: a yellow, roof-less pitfall trap. These traps would provide the best of both options: they are deeper than a pan trap (likely a good for collecting some Arthropods), but are yellow and without a cover (other features that are good for capturing many flying insects). These are actually very similar to a design that is already being used with a long-term ecological monitoring program in Greenland. We think they have it right**.
A yellow pitfall trap - the kind used in Greenland, and the one we recommend for future monitoring in Canada’s Arctic.
In sum, this work is really a “methodological” study, which when viewed narrowly may not be that sexy. However, we are optimistic that this work will help guide future ecological monitoring programs in the north. We are faced with increased pressures on our environment, and a pressing need to effectively track these effects on our biodiversity. This requires sound methods that are feasible and provide us with a true picture of faunal diversity and community structure.
It looks to me like we can capture northern beetles and spiders quite efficiently with, um, yellow plastic beer cups. Cheers to that!
Reference
Ernst, C, S. Loboda and CM Buddle. 2015. Capturing Northern Biodiversity: diversity of arctic, subarctic and northern boreal beetles and spiders are affected by trap type and habitat. Insect Conservation and Diversity DOI: 10.1111/icad.12143
——
* The paper isn’t open access. One of the goals of this blog post is to share the results of this work even if everyone can’t access the paper directly. If you want a copy of the paper, please let me know and I’ll be happy to send it to you. I’m afraid I can’t publish all of our work in open access journals because I don’t have enough $ to afford high quality OA journals.
** The big caveat here is that a proper quantitative study that compares pan and and pitfall traps to the “yellow roof-less pitfall” traps is required. We believe it will be the best design, but belief does need to be backed up with data. Unfortunately these kind of trap-comparison papers aren’t usually high on the priority list.
When asked “how are you”, many people give a default answer of “fine”. Although that may be true a lot of the time, I worry that not everyone is “fine” all of the time. In my experience, this is especially true at this time of year: the late autumn can be tough on a lot of people, as the semester is no longer new and exciting, and the dark days of November* are ahead.
I’m involved with a lot of initiatives on campus around ‘wellness’ of our community, from mental and physical health, through to trying to best understand our campus resources, and think about ways we can be proactive around well-being. Part of my goal is to increase awareness of services and resources available to everyone, and to ‘check in’ with people as we enter a difficult time of year.
I decided to write a letter to our community, but a member of our communications team suggested a video message may also help to increase awareness. So, here’s the video. It’s low-tech and done without a script, but perhaps the message is relevant to your own community. Please share if that’s the case.
—-
*I personally find November rather tough. A few years ago my amazing and insightful wife suggested I take a photo every day in November, to explore the beauty that the month has to offer. That helped me a lot, and I’m immensely grateful for her suggestion to see beyond the dreariness to embrace colour and texture. Honestly, I think that project helped me with my own mental health, and I no longer dread November to the same degree.
I recently asked a geologist* to come to speak to my field biology class. The course is about the “St Lawrence Lowlands“, and throughout the term we visit farms, forests, lakes and streams, and we do natural history research.
Why then, do I have a geologist come and speak to us?
A result of glacial till: it’s now supporting biodiversity.
Ecology is built upon geology. This may seem obvious, but requires a deeper discussion: after hearing this guest lecture year after year, I no longer see my local landscape as some farm fields, patches of forests, and some big bodies of water**. I see lands and waters shaped by a history before our time. The local landscape is a product of past geological events. We have farm fields around the Montérégie because the Champlain Sea deposited its sediments and after it departed; what remained is a flat expanse, perfect for farming. As the sea departed, it left behind remnants of beaches still visible today, as the Plateau district of Montreal, or where apple orchards grow next to Mont St Hilaire. We have some slight elevation here and there because of sandy deposits left by the departure of the last great glacier that covered our land in the very recent past. That’s where we find great white pines, stretching up above the canopies of the deciduous trees. We have Mount Rigaud because of processes hundreds of millions of years ago: an igneous intrusion that happened long, long before the age of dinosaurs. More recent igneous intrusions created the Lachine rapids, historically important as this became a key place where First Nations people, and later Europeans, set up camp along their journey up or down the big river. This was the one of the birthplaces of Montreal.
Our landscape, and the ecology of our landscape, is built upon slow but incredible processes, and I think biologists don’t consider those processes as dynamic forces that are constantly influencing our current view of the world. Ecologists often think of time in scales of decades or centuries, and we spend considerable time looking at time frames that resonate with our own life spans (in contrast, evolutionary biologists and taxonomists look much further back, and are accustomed to time frames of ‘millions of years’. I think We need to meet in the middle a little more).
As field biologists, knowing the origin of those big rocks in the forest matters a great deal: glacial till from the past creates habitats today. Moss creeps on these ancient boulders; centipedes and spiders crawl underneath. Their ephemeral life depends on much longer time frames. It’s hard to imagine how to consider discussion land management or wildlife conservation in the region without appreciating how past geological events can either help or hinder the process. There’s a geological reason why soil development is slow in some parts of our local ecosystems; why the land may be rocky, and why it’s well-drained in some areas, and wet in others. This affects long-term planning around wildlife preserves, or housing developments. There’s good reason why Mont St Hilaire is a biosphere reserve, and how it’s flora and fauna will be different that what we find elsewhere in the St Lawrence Lowlands.
Hiking at Mont St Hilaire: there are so many reasons why it’s a special place, including geology.
The longer I spend living here and learning about my region’s natural history, the more I recognize the value of some knowledge about geology, and this is why I have a geologist give a guest lecture. The students also tell me, year after year, that they appreciate and value this perspective, and their understanding of this part of the world is enriched by a deeper discussion about ‘why’ the St Lawrence Lowlands exists as it does.
How often do ecological classes include discussion about geology? Perhaps not often enough.
—-
*the geologist in question is Dr. George McCourt, who teaches often in the McGill School of Environment. I am immensely thankful for him taking time to teach us about his passion.
**when I commute to work, this is what I see: forests, field and lakes. Others in the St Lawrence Lowlands will have a different story, perhaps one that involves highrises and concrete.